
Profaza I mejozy to niezwykle złożony i kluczowy etap podziału komórkowego, który decyduje o zmienności genetycznej organizmów. Ten artykuł szczegółowo wyjaśni wszystkie stadia profazy I, od leptotenu po diakinezę, ukazując ich znaczenie dla dziedziczenia i ewolucji.
Profaza I mejozy – fundament zmienności genetycznej
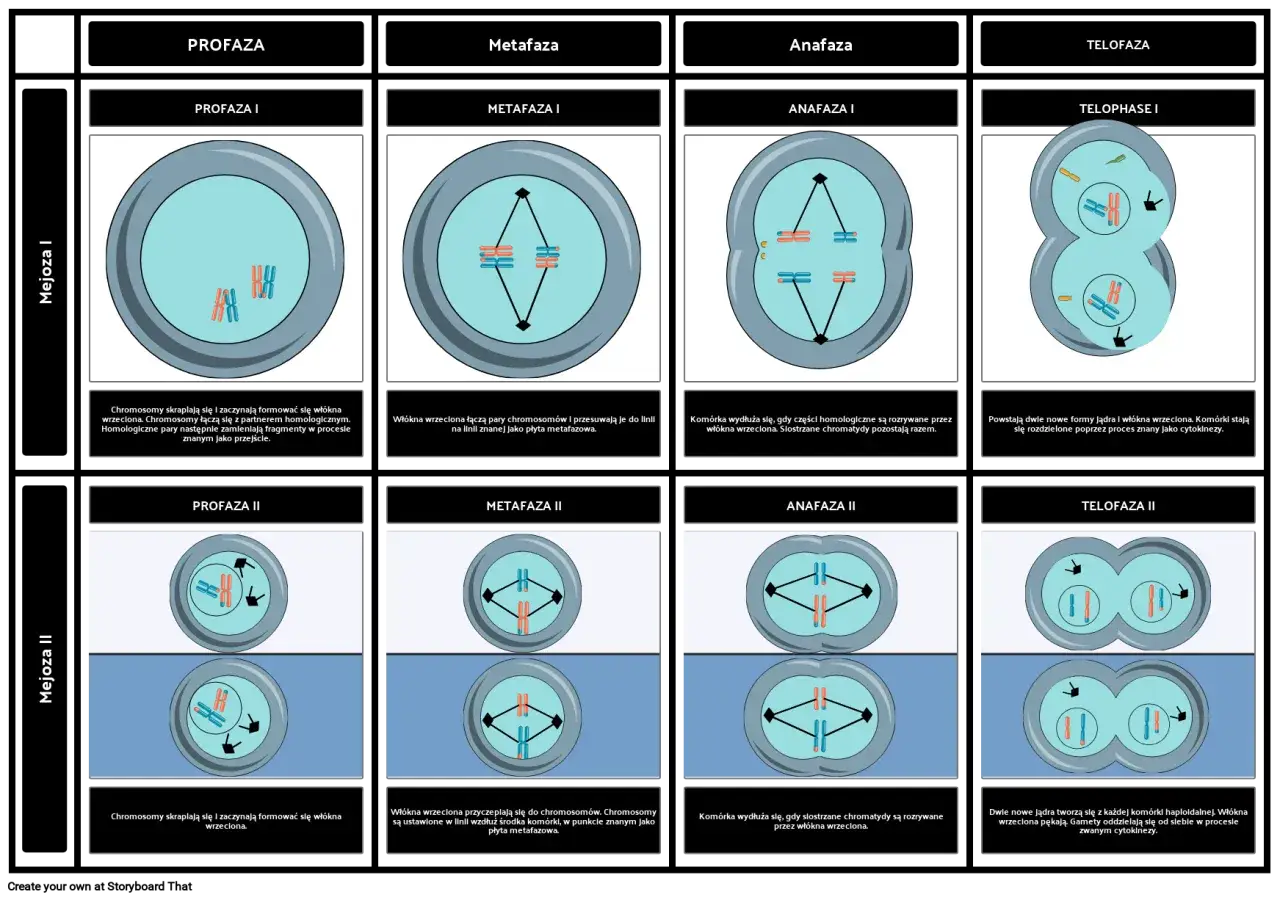
- Profaza I jest pierwszym i najbardziej złożonym etapem mejozy I, kluczowym dla rekombinacji genetycznej.
- Dzieli się na pięć charakterystycznych stadiów: leptoten, zygoten, pachyten, diploten i diakinezę.
- W zygotenie chromosomy homologiczne parują się, tworząc biwalenty (tetrady chromatyd).
- W pachytenie zachodzi crossing-over, czyli wymiana fragmentów materiału genetycznego między chromatydami niesiostrzanymi.
- Crossing-over prowadzi do powstania nowych kombinacji alleli, co jest fundamentalne dla ewolucji i adaptacji gatunków.
- Diploten charakteryzuje się rozdzielaniem chromosomów, które pozostają połączone w miejscach chiazm.
Profaza I mejozy – dlaczego ten etap jest kluczem do zmienności genetycznej?
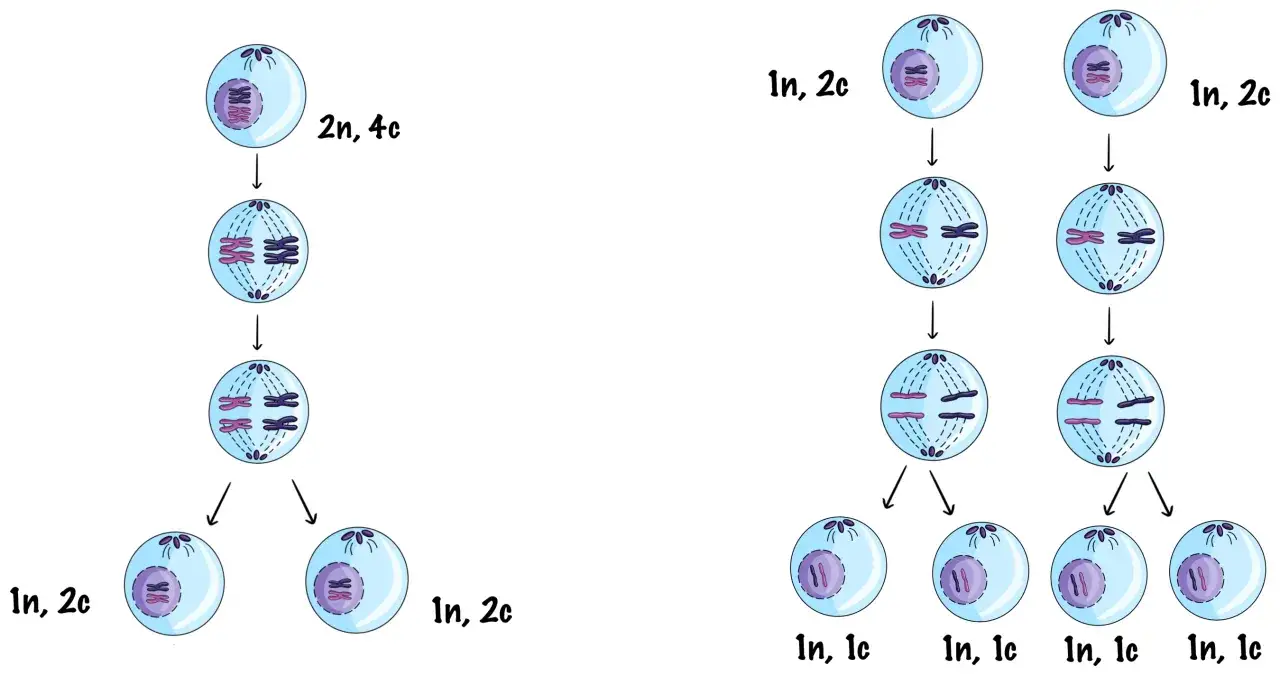
Profaza I mejozy stanowi serce całego procesu podziału mejotycznego, będąc etapem o fundamentalnym znaczeniu dla zapewnienia zmienności genetycznej organizmów. To właśnie tutaj zachodzą unikalne procesy, które odróżniają mejozę od mitozy, a które są absolutnie kluczowe dla mechanizmów dziedziczenia. Bez tych zdarzeń, potomstwo nie różniłoby się od rodziców, a ewolucja w znanym nam kształcie nie mogłaby postępować.
Rola i znaczenie profazy I w kontekście całego podziału mejotycznego
Profaza I jest bez wątpienia najbardziej złożonym i krytycznym etapem mejozy I. To w jej trakcie dochodzi do dwóch fundamentalnych zdarzeń: koniugacji chromosomów homologicznych oraz crossing-over. Oba te procesy są nie tylko niezbędne do prawidłowego przebiegu dalszych faz mejozy, ale przede wszystkim gwarantują, że każda komórka potomna otrzyma unikalny zestaw genów. Bez skutecznego sparowania chromosomów i wymiany między nimi materiału genetycznego, mejoza nie mogłaby efektywnie pełnić swojej roli w rozmnażaniu płciowym, jakim jest produkcja zróżnicowanych gamet.
Czym profaza I różni się od profazy w mitozie?
Profaza I mejozy i profaza mitozy, choć obie są początkowymi etapami podziału jądra komórkowego, różnią się diametralnie pod względem zachodzących w nich procesów. Najbardziej uderzającą różnicą jest fakt, że w profazie I mejozy dochodzi do parowania chromosomów homologicznych (koniugacja), czyli chromosomów pochodzących od matki i ojca, które niosą geny dla tych samych cech. W profazie mitozy chromosomy homologiczne nie parują się; każdy chromosom zachowuje się niezależnie.
Kolejną kluczową cechą profazy I jest tworzenie biwalentów (nazywanych również tetrady chromatyd), które są strukturami złożonymi z dwóch sparowanych chromosomów homologicznych, a więc z czterech chromatyd. W profazie mitozy takie struktury nie powstają. Co najważniejsze, w profazie I mejozy zachodzi crossing-over proces wymiany fragmentów materiału genetycznego między chromatydami niesiostrzanymi chromosomów homologicznych. To właśnie crossing-over jest głównym źródłem rekombinacji genetycznej. W profazie mitozy crossing-over nie występuje. Te unikalne zjawiska profazy I są gwarancją genetycznej różnorodności potomstwa.
Podróż przez pięć stadiów profazy I: szczegółowy przewodnik krok po kroku
Profaza I mejozy jest procesem złożonym, który przebiega sekwencyjnie przez pięć odrębnych stadiów. Każde z nich charakteryzuje się specyficznymi zmianami w jądrze komórkowym, które kumulują się, prowadząc do kluczowych zdarzeń rekombinacyjnych i przygotowując komórkę do dalszych etapów mejozy.
Leptoten: Jak z chromatyny wyłaniają się chromosomy?
Leptoten to początek profazy I, moment, w którym materiał genetyczny, dotychczas rozproszony w jądrze w postaci luźnej chromatyny, zaczyna ulegać kondensacji. Chromosomy stają się stopniowo widoczne pod mikroskopem jako cienkie, długie nici. Ważne jest, aby pamiętać, że każdy z tych widocznych chromosomów już w tym stadium składa się z dwóch identycznych siostrzanych chromatyd, połączonych centromerem. Choć chromatydy są już obecne, nie są one jeszcze wyraźnie rozdzielone wizualnie.
Zygoten: Na czym polega "parowanie" chromosomów homologicznych i tworzenie biwalentów?
Zygoten to stadium, w którym rozpoczyna się kluczowy proces koniugacji, znany również jako synapsis. Polega on na precyzyjnym zbliżaniu się do siebie i łączeniu chromosomów homologicznych pary chromosomów, z których jeden odziedziczyliśmy po matce, a drugi po ojcu. Połączenie to jest bardzo ścisłe i odbywa się dzięki specjalnej strukturze białkowej, zwanej kompleksem synaptonemalnym. Wynikiem koniugacji jest powstanie biwalentu, czyli struktury składającej się z dwóch połączonych chromosomów homologicznych. Ponieważ każdy chromosom w tym momencie składa się z dwóch chromatyd, biwalent zawiera łącznie cztery chromatydy, dlatego często nazywany jest także tetrady chromatyd.
Pachyten: Serce rekombinacji, czyli jak działa mechanizm crossing-over?
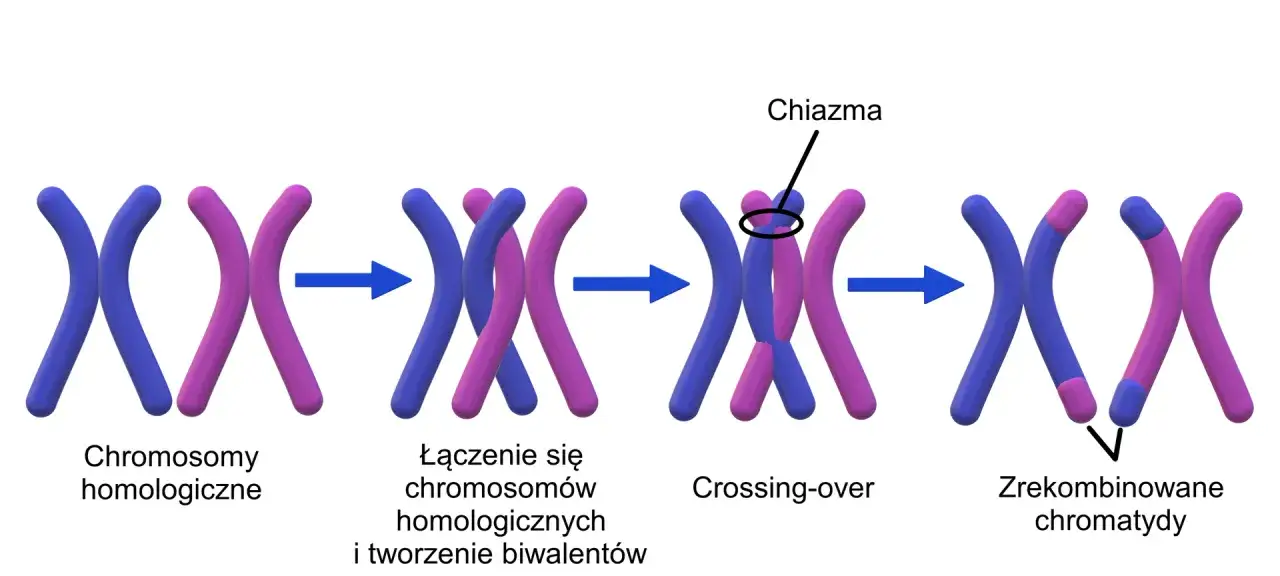
Pachyten jest bez wątpienia najbardziej doniosłym stadium profazy I, ponieważ to właśnie w nim zachodzi fundamentalny proces crossing-over. Jest to mechanizm, podczas którego dochodzi do wymiany fragmentów materiału genetycznego pomiędzy chromatydami niesiostrzanymi należącymi do chromosomów homologicznych. Wyobraźmy sobie, że chromatydy te "wymieniają się" fragmentami DNA. Miejsca, w których doszło do tej wymiany, są fizycznie widoczne później jako chiazmy. Crossing-over jest kluczowe dla zmienności genetycznej, ponieważ prowadzi do powstania nowych kombinacji alleli na chromosomach, co jest podstawą dziedziczenia cech w sposób inny niż u rodziców.
Diploten: Co się dzieje po wymianie genów i czym są tajemnicze chiazmy?
W stadium diplotenu obserwujemy, że chromosomy homologiczne w obrębie biwalentów zaczynają się od siebie odsuwać i rozdzielać. Jednakże, co niezwykle istotne, nie rozchodzą się one całkowicie. Pozostają połączone w tych specyficznych miejscach, gdzie wcześniej zaszła wymiana materiału genetycznego, czyli właśnie w chiazmach. Chiazmy są zatem widocznym, fizycznym dowodem na to, że crossing-over faktycznie miał miejsce, i odgrywają one kluczową rolę w utrzymaniu biwalentu w całości aż do momentu rozdzielenia chromosomów homologicznych w późniejszej fazie mejozy.
Diakineza: Finalne przygotowania do dalszego podziału komórki
Diakineza stanowi ostatnie stadium profazy I, charakteryzujące się maksymalną kondensacją chromosomów. Stają się one grubsze i krótsze, co ułatwia ich późniejsze rozdzielenie. W tym czasie obserwuje się również zjawisko terminalizacji chiazm chiazmy, które były początkowo szeroko rozstawione, przesuwają się w kierunku końców nici chromosomowych. Jednocześnie, otoczka jądrowa rozpada się, a jąderko zanika. Wszystkie te zmiany przygotowują komórkę do przejścia w kolejną fazę mejozy I metafazę I.
Crossing-over: Jak wymiana genów kształtuje życie, które znamy?
Proces crossing-over jest jednym z najbardziej fundamentalnych zjawisk biologicznych, które kształtuje życie na Ziemi w sposób, jaki znamy. To właśnie dzięki tej subtelnej, ale potężnej wymianie fragmentów DNA pomiędzy chromosomami rozmnażającymi się płciowo gatunkami, możliwe jest ciągłe tworzenie się nowych kombinacji cech i adaptacja do nieustannie zmieniającego się środowiska.
Mechanizm molekularny: Co dokładnie dzieje się na poziomie DNA?
Na poziomie molekularnym, crossing-over jest złożonym procesem biochemicznym. Rozpoczyna się od precyzyjnych pęknięć w podwójnej helisie DNA obu chromatyd niesiostrzanych, które następnie są naprawiane przez specjalne enzymy. W trakcie tej naprawy dochodzi do wymiany fragmentów DNA pomiędzy nimi. Kompleks synaptonemalny, który utrzymuje chromosomy homologiczne blisko siebie przez cały okres profazy I, odgrywa kluczową rolę w zapewnieniu, że wymiana ta zachodzi między odpowiednimi fragmentami chromosomów. Jest to niezwykle precyzyjny mechanizm, który gwarantuje, że na końcu procesu powstają nowe, unikalne sekwencje genetyczne.
Znaczenie biologiczne: Dlaczego rekombinacja genetyczna jest fundamentem ewolucji?
Biologiczne znaczenie crossing-over jest nie do przecenienia. Tworzenie nowych kombinacji alleli czyli różnych wersji tego samego genu w obrębie chromosomów znacząco zwiększa zmienność genetyczną w populacji. Ta zmienność jest kluczowym "surowcem" dla działania doboru naturalnego. Dzięki niej osobniki posiadające korzystne kombinacje cech mają większe szanse na przeżycie i reprodukcję w danym środowisku. W dłuższej perspektywie, ciągła rekombinacja genetyczna umożliwia gatunkom adaptację do zmieniających się warunków środowiskowych i jest jednym z głównych motorów napędowych ewolucji, prowadząc do powstawania nowych gatunków i różnorodności życia na Ziemi.
Wizualizacja profazy I: Jak wyglądają chromosomy na każdym etapie?
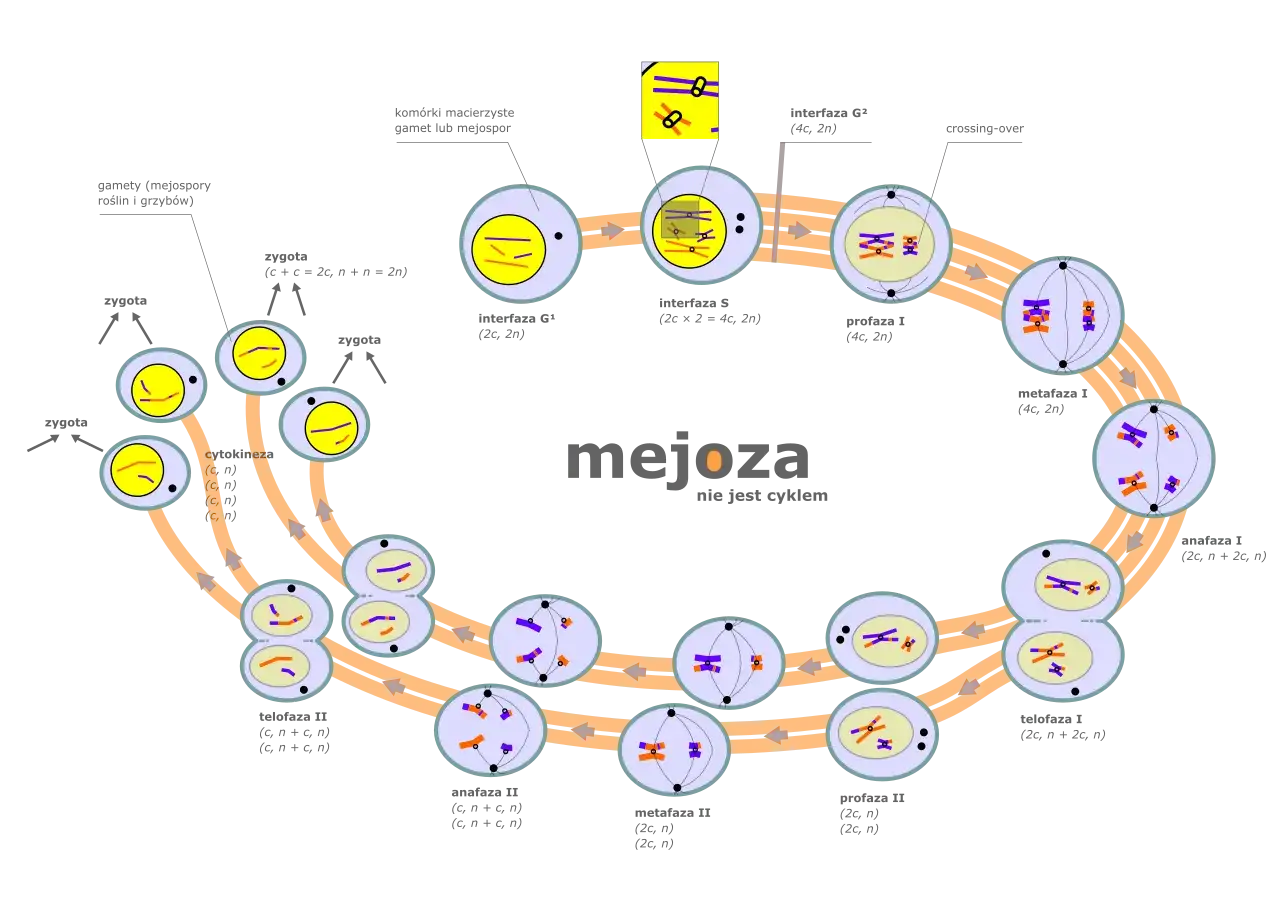
Zrozumienie złożonych procesów zachodzących w profazie I mejozy jest znacznie łatwiejsze, gdy możemy je sobie zwizualizować. Obserwacja zmian morfologicznych chromosomów na poszczególnych etapach pozwala nam śledzić postęp tego fascynującego procesu i docenić jego znaczenie.
Od cienkich nici do skondensowanych biwalentów – graficzne przedstawienie zmian
W stadium leptotenu chromosomy są początkowo widoczne jako bardzo cienkie, długie nici chromatyny, które dopiero zaczynają się kondensować. W zygotenie, te nici zaczynają się parować, tworząc struktury dwurzędowe biwalenty. W pachytenie biwalenty stają się bardziej zwarte, a na schematach można już dostrzec miejsca potencjalnego crossing-over. W diplotenie biwalenty zaczynają się rozdzielać, ale pozostają połączone w chiazmach, co nadaje im charakterystyczny, krzyżowy wygląd. Wreszcie, w diakinezie chromosomy osiągają maksymalną kondensację, są grube i krótkie, a chiazmy przesuwają się ku końcom, przygotowując je do metafazy I.
Obraz mikroskopowy: Czy można zobaczyć profazę I "na żywo"?
Obserwacja profazy I pod mikroskopem jest możliwa, choć wymaga wprawy i odpowiednich technik. Poszczególne stadia można rozpoznać na podstawie morfologii chromosomów ich długości, grubości oraz obecności chiazm. Do uwidocznienia tych struktur często stosuje się specjalne barwniki, które zwiększają kontrast i pozwalają na lepszą wizualizację. Obserwacja "na żywo" jest jednak trudna ze względu na dynamikę procesów zachodzących w komórce. Zazwyczaj analizuje się preparaty utrwalone, gdzie można dokładnie zbadać strukturę chromosomów w poszczególnych fazach profazy I. Uzyskanie wyraźnych obrazów mikroskopowych jest kluczowe dla potwierdzania przebiegu mejozy i identyfikacji ewentualnych nieprawidłowości.
Najczęstsze błędy i nieprawidłowości w przebiegu profazy I – jakie są ich konsekwencje?
Prawidłowy przebieg profazy I jest absolutnie kluczowy nie tylko dla zdrowia reprodukcyjnego jednostki, ale także dla stabilności genetycznej populacji. Wszelkie błędy i nieprawidłowości na tym etapie mogą prowadzić do poważnych konsekwencji, wpływając na płodność, zdrowie potomstwa, a nawet na proces ewolucji gatunku.
Problem z parowaniem chromosomów i jego wpływ na płodność
Jednym z najpoważniejszych błędów, który może wystąpić podczas profazy I, jest tzw. non-disjunction, czyli brak prawidłowego rozchodzenia się chromosomów homologicznych. Jeśli chromosomy homologiczne nie sparują się poprawnie lub nie zostaną prawidłowo utrzymane w biwalentach, mogą nie rozdzielić się właściwie podczas późniejszych faz mejozy. Skutkuje to powstawaniem gamet (plemników lub komórek jajowych) z nieprawidłową liczbą chromosomów, czyli z tzw. aneuploidią. Na przykład, jeśli dojdzie do non-disjunction chromosomu 21, zapłodnienie takiej komórki może prowadzić do rozwoju zespołu Downa. Aneuploidia często jest przyczyną niepłodności, poronień samoistnych lub narodzin dzieci z poważnymi wadami rozwojowymi.
Przeczytaj również: Biologia: Czym jest genetyka i jak wpływa na dziedziczenie cech
Gdy crossing-over nie zachodzi: skutki braku rekombinacji
Zaburzenia w procesie crossing-over, w tym jego całkowity brak, mają równie dalekosiężne konsekwencje. Brak rekombinacji genetycznej oznacza, że potomstwo będzie dziedziczyć geny w dokładnie takich samych kombinacjach, w jakich występowały one u rodziców. Prowadzi to do znaczącego zmniejszenia zmienności genetycznej w populacji. W dłuższej perspektywie, taka jednorodność genetyczna osłabia zdolność populacji do adaptacji do zmieniających się warunków środowiskowych, czyniąc ją bardziej podatną na choroby czy zmiany klimatyczne. Ponadto, nieprawidłowo utworzone lub niestabilne chiazmy mogą również zakłócać prawidłową segregację chromosomów w anafazie I, co ponownie może prowadzić do powstawania gamet z nieprawidłową liczbą chromosomów i problemów z płodnością.